







































mitochondrion
 The mitochondrion (pl. mitochondria) is the 'power house of the eukaryotic cell, performing oxidative phosporylation. Mitochondria have two internal, membrane-bound spaces, unlike chloroplasts, which have three internal spaces. The outer mitochodrial membrane is similar in constitution to the eukaryotic cell’s plasma membrane, while the inner membrane is similar in chemical composition to bacterial membranes. This difference is one of several lines of evidence for the serial endosymbiotic origin of mitochondria as phagocytozed purple bacteria.
The mitochondrion (pl. mitochondria) is the 'power house of the eukaryotic cell, performing oxidative phosporylation. Mitochondria have two internal, membrane-bound spaces, unlike chloroplasts, which have three internal spaces. The outer mitochodrial membrane is similar in constitution to the eukaryotic cell’s plasma membrane, while the inner membrane is similar in chemical composition to bacterial membranes. This difference is one of several lines of evidence for the serial endosymbiotic origin of mitochondria as phagocytozed purple bacteria.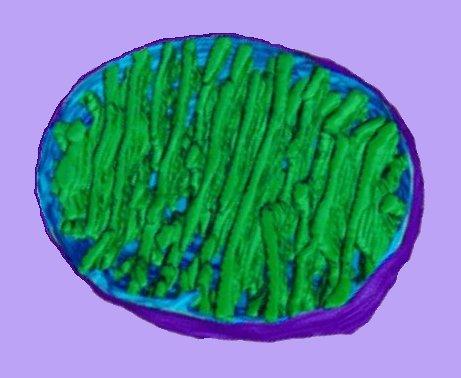{kind=link}
Above left – click to enlarge : simplified diagram of a mitochondrion showing
1. intermembranous space between inner and outer membranes : protons of chemiosmotic gradient : phosphorylation of nucleotides,
2. matrix : mtDNA, ribosomes : Krebs tricarboxylic acid cycle, part of urea cycle.
3. christae (expand surface area of the inner membrane)
4. junction between membranes
5. inner membrane : translocase (TIM, Tim complex) : cytochromes, electron transport chain, oxidative phosporylation, ATP synthase.
6. outer membrane : translocase (TOM, Tom complex) : oxidation of epinephrine (adrenaline), degradation of tryptophan, and elongation of fatty acids.
7. organelle surrounded by cytosol : ribosomes and ribozymes of rER : glyoxylate shunt, part of urea cycle.
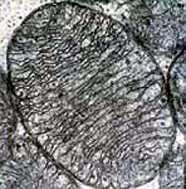
Left - transmission electron micrograph (tem) of mitochondrion.
{kind=link}

The image at right (click image to enlarge) is based on reconstruction of serial tem slices through a mitochondrion. The outer membrane (violet) surrounds the organelle. The inner membrane (pale blue) is contiguous, at membrane junctions (pale blue connecting to green at lower center), with the inner membrane that forms the walls of cristae (green).
The matrix – a soup factory – lies between the cristae, and contains mitochondrial DNA and components of intermediary metabolism. image - mitochondrion cut : tour mitochondrion :
{kind=link}
{kind=link}
The outer and inner membranes are composed of phospolipid bilayers studded with proteins, much like the cell membrane. However, the composition of the inner and outer membranes is very different.
The inner mitochondrial membrane contains more than 100 different polypeptides. The protein to phospholipid ratio is very high – more than 3:1 by weight, having about 1 protein for 15 phospholipids. The inner membrane is also rich in an unusual phospholipid, cardiolipin, which is usually characteristic of bacterial plasma membranes, and which renders the membrane virtually impermeable. This composition, along with other evidence, has led to the assumption that the inner membrane is derived from endosymbiotic prokaryotes. The endosymbiotic theory of eukaryotic evolution is now widely accepted. Active transport of large molecules across the inner membrane is effected by translocases in the Tim complex. Proteins must posssess a NH2 cleavable signal sequence in order to reach the matrix via the Tom and Tim complexes [more].
In contrast, the outer membrane, which encloses the entire mitochondrion, is similar in composition to the cell's plasma membrane and comprises about 50% phospolipids by weight and contains a variety of enzymes. The outer membrane contains integral porin proteins, which contain a channel (2-3nm) that permits passage of molecules up to 10,000 daltons. Larger molecules cross the membrane by active transport through the translocase (Tom complex). Membranous enzymes carry out activities such as the oxidation of epinephrine (adrenaline), the degradation of tryptophan, and the elongation of fatty acids.
'Research over the last decade has extended the prevailing view of mitochondria to include functions well beyond the critical bioenergetics role in supplying ATP. It is now recognized that mitochondria play a crucial role in cell signaling events, inter-organelle communication, aging, many diseases, cell proliferation and cell death. Apoptotic signal transmission to the mitochondria results in the efflux of a number of potential apoptotic regulators to the cytosol that trigger caspase activation and lead to cell destruction. Accumulating evidence indicates that the voltage-dependent anion channel (VDAC) is involved in this release of proteins via the outer mitochondrial membrane. VDAC in the outer mitochondrial membrane is in a crucial position in the cell, forming the main interface between the mitochondrial and the cellular metabolisms. VDAC has been recognized as a key protein in mitochondria-mediated apoptosis since it is the proposed target for the pro- and anti-apoptotic Bcl2-family of proteins and due to its function in the release of apoptotic proteins located in the inter-membranal space (AIF, Endo-G, Smac, cytochrome c). The diameter of the VDAC pore is only about 2.6-3 nm, which is insufficient for passage of a folded protein like cytochrome c. New work suggests pore formation by homo-oligomers of VDAC or hetero-oligomers composed of VDAC and pro-apoptotic proteins such as Bax or Bak.' Shoshan-Barmatz V, Israelson A, Brdiczka D, Sheu SS. The voltage-dependent anion channel (VDAC): function in intracellular signalling, cell life and cell death. Curr Pharm Des. 2006;12(18):2249-70.
The plant chloroplast is the site of photosynthesis : animation - chloroplast : tour the chloroplast : Virtual Cell Textbook - Cell Biology
{kind=link}
Џ beautiful Flash 8 animation - Inner Life of the Cell, which shows mitochondrion, and Interpretation: Inner Life of the Cell Џ
• A • adhesion • C • cell membranes • cellular adhesion molecules • cellular signal transduction • centrioles • chemotaxis • chloroplast • cilia & flagella • communication • concentration gradients • cytokine receptors • cytoplasm • cytoskeleton • E • energy transducers • endoplasmic reticulum • endosomes • exosome • F • flagella & cilia • G • Golgi apparatus • GPCRs • H • hormones • I • ion channels • L • lysosome • M • meiosis • microtubules • mitosis • mitochondrion • N • Nitric Oxide • neurotransmission • neuronal interconnections • nuclear membrane • nuclear pore • P • pinocytosis • proteasome • pumps • R • receptor proteins • receptor-mediated endocytosis • S • second messengers • signaling gradients • signal transduction • spindle • structure • T • transport • two-component systems • V • vacuole • vesicle •