







































chemotaxis
Chemotaxis is a signal transduction system that confers to the ability of cells to respond to spatial and temporal chemical gradients. In prokaryotes and unicellular eukaryotes, chemotaxis controls motile behavior. In multicellular eukaryotes, chemotaxis plays an important role in determining the migration of cells in many biological processes including immune response, development, embryogenesis, wound healing, and angiogenesis (1, 2, 3, 4). During embryogenesis, cells respond to chemotactic stimuli, enabling the organization of tissues and organs, and neuronal migration (5, 6, 7). An elaborate network of chemoattractants directs leukocytes to their correct locations and facilitates cell-cell interactions in the immune system. Chemotaxis is central to wound healing, and operates in disease states such as metastasis and atherosclerosis (8, 9, 10, 11, 12).
Even though the motility apparatus differs among organisms, the general mechanism of control of chemotaxis is conserved throughout all prokaryotic bacteria and archaea. Central to chemotactic control is the two-component system in which phosphorylation of a response regulator reflects phosphorylation of a histidine autokinase that senses environmental parameters (117). This is the commonest mode of signal transduction system in bacteria, and the two component system controls diverse processes such as gene expression, sporulation, and chemotaxis.
Prokaryotic chemotaxis proteins comprise four groups—a signal recognition and transduction group, an excitation group, an adaptation group, and a signal removal group (to dephosphorylate CheY-P). The signal recognition and transduction group includes the receptors and ligand binding proteins, which are capable of binding effectors outside the cell; a few receptors, however, are cytoplasmic: (image general chemotaxis model : Table proteins in chemotaxis ). Prokaryotes react as point sensors, determining static gradients by temporal sampling.
The mechanism of chemotaxis in eukaryotic cells is different than that in prokaryotes, though eukaryotic cells also respond to chemical, signaling gradients. By virtue of their greater size, eukaryotes are able to detect concentration gradients along their plasma membranes where receptor occupancy differences provide input for discrepancy sensing of gradients. In response to chemoattractant gradients, the imbalance between activator action and inactivator action results in a spatially oriented persistent signaling. Most eukaryotic cells employ GPCRs to detect these chemical signals, so signaling is amplified by a substrate supply-based positive feedback that acts through small G-proteins. This amplification is activated only in the continuous presence of the external signal gradient, thus providing the mechanism for sensitivity to gradient alterations. Other membrane receptor include amino acids, insulin, and vasoactive peptides. Chemotactic receptors are triggered by formyl peptides, chemokines (α, CXC; β, CC; δ, CX3C; γ, C), and leukotrienes (autocrine and paracrine eicosanoid lipid mediators derived from arachidonic acid by 5-lipoxygenase).
Following signal detection by a GPCR, a locally acting activator (PI3-kinase) and a globally acting inactivator (PTEN or a similar phosphatase) are coordinately controlled by the G-protein activation [1]. The signaling system adapts to spatially homogeneous changes in the chemoattractant. In chemoattractant gradients, an imbalance between the action of the activator and the inactivator results in a spatially oriented persistent signaling, amplified by a substrate supply-based positive feedback acting through small G-proteins. The amplification is activated only in a continuous presence of the external signal gradient, thus providing the mechanism for sensitivity to gradient alterations.
During the course of evolution, eukaryotic cilia have been adapted to function as mechanoreceptors and chemoreceptors. Eukaryotic cilia and flagella differ from the flagella of eubacteria, which differ in turn from the flagella of archaebacteria. Organisms such as Amoebae and Tetrahymena are able to creep or swim, utilizing the cytoskeleton/cilia/flagella. The slime mold, Dictyostelium discoideum commences gradient-sensing with modification of the phospholipid membrane at the cell's leading edge. The modification includes the recruitment of various proteins to the membrane, initiating a process that culminates in pseudopod extension in the direction of the chemoattractant. Cells exposed to shallow chemoattractant gradients respond with strong accumulation of the enzyme phosphatidylinositol 3-kinase (PI3-K, PI3K) and its D3-phosphoinositide product (PIP3) at the plasma membrane exposed to the highest chemoattractant concentration, whereas PIP3-degrading enzyme PTEN and its product phosphatidylinositol bisphosphate (PIP2) localize in a complementary pattern [Gamba]. This early symmetry-breaking event is a mandatory step for directed cell movement elicited by chemoattractants. It has been suggested that 'directional sensing is the consequence of a phase-ordering process mediated by phosphoinositide diffusion and driven by the distribution of chemotactic signal'.
When starved, D. discoidem cells differentiate, polarize, and migrate directionally toward secreted 3',5'-cyclic adenosine monophosphate (cAMP). The cAMP is detected by four GPCRs, designated cAR1–cAR4, which are coupled to a single heterotrimeric G-protein (19). Mammalian leukocytes couple 20 types of chemoattractant, chemokine receptors to the same G-protein, Gi (20, 21). Mammalian systems also employ chemoattractant-elicited transient increases in phosphoinositides (PIs),1 cAMP, cGMP, inositol trisphosphate (IP3), and Ca2+, and rearrangements in the cytoskeleton (16, 22). The PI3-kinase elevates local levels of PIP3, which determines the sites of new actin-filled projections. PIP3 is an important intermediate in chemotactic signaling in D. discoideum, amoebae, and mammalian leukocytes (23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34). 'Robust control of the temporal and spatial levels of PIP3 is achieved by reciprocal regulation of PI3K and PTEN.'[2, fft]
Ras, PI3K, and TOR are known as key regulators of cellular growth, and play a critical role in regulating the actin cytoskeleton, cell polarity, and cellular movement, indicating that multiple steps in the signal transduction pathway coordinately regulate cell motility.
Chemotaxis is important early in embryologic development and in the immune system. Within multicellular organisms, specific types of cells such as those of the immune/hematologic system (granulocytes, monocytes, macrophages, lymphocytes) and specialized, independent cells (mast cells, endothelial cells, fibroblasts, and, unfortunately, tumor cells) are capable of motile responses to chemical signals. Motility and migration employ cellular adhesion molecules.
Џ beautiful Flash 8 animation - Inner Life of the Cell, which shows chemotaxis in action, and Interpretation of Inner Life of the Cell Џ
· adenylyl (adenylate) cyclase · calcium ions · cAMP-dependent protein kinase · CDKs · cyclin-dependent kinases · DAG · diacylglycerol · DNA ligases · ERKs · GPCRs · GPCR families · guanylate cyclases · guanyl cyclase · inositol triphosphate · IP3 · MAP kinases · mitogen activated protein kinases · phosphatases · phosphodiesterases · phospolipases · phosphorylation · PKA · PKC · phospholipase C-gamma · protein kinase A · protein kinase C · protein tyrosine kinases (PTKs) · receptor tyrosine kinases · second messengers · second messenger cAMP · second messenger cGMP · signal transduction · two-component systems ·
Tables Cell signaling Cell Adhesion Molecules Immune Cytokines Second Messengers Receptor Tyrosine Kinases(RTK) Second Messengers Phosphate-handling Enzymes
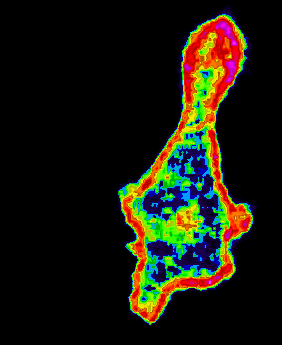
: confocal image -GFB amoeba chemotaxis : : Links to videos of Dictyostelium: Cytokinesis : Cytokinesis Defects : Chemotaxis : Development : Morphogenesis : Motility : Phagocytosis : Videos Chemotaxis : Chemoattractant receptors in chemotaxis : G-protein b-subunits in chemotaxis : PH-domain translocation with uniform stimulus : PH-domain translocation during chemotaxis : PH-domain translocation in immobilized cells : PH-domain dynamics in immobilized cells : Digitized wild-type cell in chemotaxis : Digitized clathrin heavy chain null cell in chemotaxis :
In addition to chemotaxis, cells perform:
1) chemokinesis – scanning the environment,
2) haptotaxis – generation of a chemoattractant gradient in the ECM employed in transendothelial migration and angiogenesis, and
3) necrotaxis – negative or positive response to chemoattractants released from apoptotic or necrotic cells.
• A • adhesion • C • cell membranes • cellular adhesion molecules • cellular signal transduction • centrioles • chemotaxis • chloroplast • cilia & flagella • communication • concentration gradients • cytokine receptors • cytoplasm • cytoskeleton • E • energy transducers • endoplasmic reticulum • endosomes • exosome • F • flagella & cilia • G • Golgi apparatus • GPCRs • H • hormones • I • ion channels • L • lysosome • M • meiosis • microtubules • mitosis • mitochondrion • N • Nitric Oxide • neurotransmission • neuronal interconnections • nuclear membrane • nuclear pore • P • pinocytosis • proteasome • pumps • R • receptor proteins • receptor-mediated endocytosis • S • second messengers • signaling gradients • signal transduction • spindle • structure • T • transport • two-component systems • V • vacuole • vesicle •

Even though the motility apparatus differs among organisms, the general mechanism of control of chemotaxis is conserved throughout all prokaryotic bacteria and archaea. Central to chemotactic control is the two-component system in which phosphorylation of a response regulator reflects phosphorylation of a histidine autokinase that senses environmental parameters (117). This is the commonest mode of signal transduction system in bacteria, and the two component system controls diverse processes such as gene expression, sporulation, and chemotaxis.
Prokaryotic chemotaxis proteins comprise four groups—a signal recognition and transduction group, an excitation group, an adaptation group, and a signal removal group (to dephosphorylate CheY-P). The signal recognition and transduction group includes the receptors and ligand binding proteins, which are capable of binding effectors outside the cell; a few receptors, however, are cytoplasmic: (image general chemotaxis model : Table proteins in chemotaxis ). Prokaryotes react as point sensors, determining static gradients by temporal sampling.
The mechanism of chemotaxis in eukaryotic cells is different than that in prokaryotes, though eukaryotic cells also respond to chemical, signaling gradients. By virtue of their greater size, eukaryotes are able to detect concentration gradients along their plasma membranes where receptor occupancy differences provide input for discrepancy sensing of gradients. In response to chemoattractant gradients, the imbalance between activator action and inactivator action results in a spatially oriented persistent signaling. Most eukaryotic cells employ GPCRs to detect these chemical signals, so signaling is amplified by a substrate supply-based positive feedback that acts through small G-proteins. This amplification is activated only in the continuous presence of the external signal gradient, thus providing the mechanism for sensitivity to gradient alterations. Other membrane receptor include amino acids, insulin, and vasoactive peptides. Chemotactic receptors are triggered by formyl peptides, chemokines (α, CXC; β, CC; δ, CX3C; γ, C), and leukotrienes (autocrine and paracrine eicosanoid lipid mediators derived from arachidonic acid by 5-lipoxygenase).
Following signal detection by a GPCR, a locally acting activator (PI3-kinase) and a globally acting inactivator (PTEN or a similar phosphatase) are coordinately controlled by the G-protein activation [1]. The signaling system adapts to spatially homogeneous changes in the chemoattractant. In chemoattractant gradients, an imbalance between the action of the activator and the inactivator results in a spatially oriented persistent signaling, amplified by a substrate supply-based positive feedback acting through small G-proteins. The amplification is activated only in a continuous presence of the external signal gradient, thus providing the mechanism for sensitivity to gradient alterations.
During the course of evolution, eukaryotic cilia have been adapted to function as mechanoreceptors and chemoreceptors. Eukaryotic cilia and flagella differ from the flagella of eubacteria, which differ in turn from the flagella of archaebacteria. Organisms such as Amoebae and Tetrahymena are able to creep or swim, utilizing the cytoskeleton/cilia/flagella. The slime mold, Dictyostelium discoideum commences gradient-sensing with modification of the phospholipid membrane at the cell's leading edge. The modification includes the recruitment of various proteins to the membrane, initiating a process that culminates in pseudopod extension in the direction of the chemoattractant. Cells exposed to shallow chemoattractant gradients respond with strong accumulation of the enzyme phosphatidylinositol 3-kinase (PI3-K, PI3K) and its D3-phosphoinositide product (PIP3) at the plasma membrane exposed to the highest chemoattractant concentration, whereas PIP3-degrading enzyme PTEN and its product phosphatidylinositol bisphosphate (PIP2) localize in a complementary pattern [Gamba]. This early symmetry-breaking event is a mandatory step for directed cell movement elicited by chemoattractants. It has been suggested that 'directional sensing is the consequence of a phase-ordering process mediated by phosphoinositide diffusion and driven by the distribution of chemotactic signal'.
When starved, D. discoidem cells differentiate, polarize, and migrate directionally toward secreted 3',5'-cyclic adenosine monophosphate (cAMP). The cAMP is detected by four GPCRs, designated cAR1–cAR4, which are coupled to a single heterotrimeric G-protein (19). Mammalian leukocytes couple 20 types of chemoattractant, chemokine receptors to the same G-protein, Gi (20, 21). Mammalian systems also employ chemoattractant-elicited transient increases in phosphoinositides (PIs),1 cAMP, cGMP, inositol trisphosphate (IP3), and Ca2+, and rearrangements in the cytoskeleton (16, 22). The PI3-kinase elevates local levels of PIP3, which determines the sites of new actin-filled projections. PIP3 is an important intermediate in chemotactic signaling in D. discoideum, amoebae, and mammalian leukocytes (23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34). 'Robust control of the temporal and spatial levels of PIP3 is achieved by reciprocal regulation of PI3K and PTEN.'[2, fft]
Ras, PI3K, and TOR are known as key regulators of cellular growth, and play a critical role in regulating the actin cytoskeleton, cell polarity, and cellular movement, indicating that multiple steps in the signal transduction pathway coordinately regulate cell motility.
Chemotaxis is important early in embryologic development and in the immune system. Within multicellular organisms, specific types of cells such as those of the immune/hematologic system (granulocytes, monocytes, macrophages, lymphocytes) and specialized, independent cells (mast cells, endothelial cells, fibroblasts, and, unfortunately, tumor cells) are capable of motile responses to chemical signals. Motility and migration employ cellular adhesion molecules.
Џ beautiful Flash 8 animation - Inner Life of the Cell, which shows chemotaxis in action, and Interpretation of Inner Life of the Cell Џ
· adenylyl (adenylate) cyclase · calcium ions · cAMP-dependent protein kinase · CDKs · cyclin-dependent kinases · DAG · diacylglycerol · DNA ligases · ERKs · GPCRs · GPCR families · guanylate cyclases · guanyl cyclase · inositol triphosphate · IP3 · MAP kinases · mitogen activated protein kinases · phosphatases · phosphodiesterases · phospolipases · phosphorylation · PKA · PKC · phospholipase C-gamma · protein kinase A · protein kinase C · protein tyrosine kinases (PTKs) · receptor tyrosine kinases · second messengers · second messenger cAMP · second messenger cGMP · signal transduction · two-component systems ·
Tables Cell signaling Cell Adhesion Molecules Immune Cytokines Second Messengers Receptor Tyrosine Kinases(RTK) Second Messengers Phosphate-handling Enzymes
: confocal image -GFB amoeba chemotaxis : : Links to videos of Dictyostelium: Cytokinesis : Cytokinesis Defects : Chemotaxis : Development : Morphogenesis : Motility : Phagocytosis : Videos Chemotaxis : Chemoattractant receptors in chemotaxis : G-protein b-subunits in chemotaxis : PH-domain translocation with uniform stimulus : PH-domain translocation during chemotaxis : PH-domain translocation in immobilized cells : PH-domain dynamics in immobilized cells : Digitized wild-type cell in chemotaxis : Digitized clathrin heavy chain null cell in chemotaxis :
{kind=link}
In addition to chemotaxis, cells perform:
1) chemokinesis – scanning the environment,
2) haptotaxis – generation of a chemoattractant gradient in the ECM employed in transendothelial migration and angiogenesis, and
3) necrotaxis – negative or positive response to chemoattractants released from apoptotic or necrotic cells.
• A • adhesion • C • cell membranes • cellular adhesion molecules • cellular signal transduction • centrioles • chemotaxis • chloroplast • cilia & flagella • communication • concentration gradients • cytokine receptors • cytoplasm • cytoskeleton • E • energy transducers • endoplasmic reticulum • endosomes • exosome • F • flagella & cilia • G • Golgi apparatus • GPCRs • H • hormones • I • ion channels • L • lysosome • M • meiosis • microtubules • mitosis • mitochondrion • N • Nitric Oxide • neurotransmission • neuronal interconnections • nuclear membrane • nuclear pore • P • pinocytosis • proteasome • pumps • R • receptor proteins • receptor-mediated endocytosis • S • second messengers • signaling gradients • signal transduction • spindle • structure • T • transport • two-component systems • V • vacuole • vesicle •