







































mitosis
Mitosis replicates somatic cells.
One cell division from single diploid parent cell produces two diploid daughter cells:
1. prophase
2. metaphase
3. anaphase
4. telophase
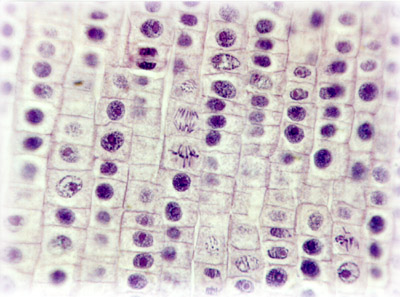
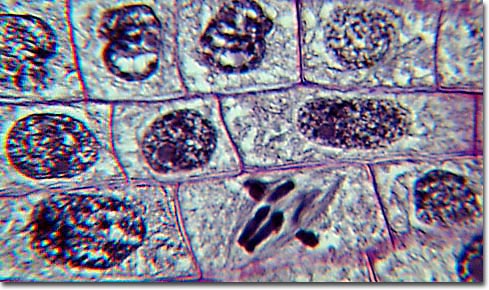
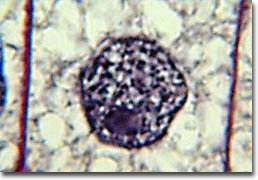
(diagram Stages of Mitosis : images : micrograph - mitotic cells : labelled micrograph : micrograph prophase metaphase anaphase : micrograph anaphase : micrograph telophase : micrograph, early prophase, late prophase, metaphase, early anaphase, late anaphase, telophase, daughter cells, resting : Movie illustration of mitosis : Mito Movie)
Mitosis allows cell nuclei to split, providing each daughter cell with a complete set of chromosomes during cellular division, while cytokinesis is division of the cytoplasm. Before replicating cells leave interphase, they pass through the synthesis or S phase in which each chromosome is duplicated and condensed to form two sister chromatids joined at the centromere. Centromeres contain a specific DNA sequence, and are crucial to segregation of the daughter chromatids during mitosis.
Mitogens, or somatomedins, are molecules that stimulates a cell to divide. Most mitogens are proteins, and they stimulate signal transduction pathways that utilize mitogen activated protein kinases. Mitogens include cytokines, growth factors, hormones, neurotransmitters, cellular stress proteins, and cell adhesion ligands.
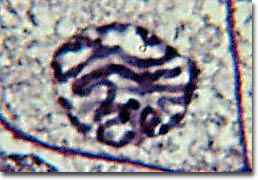
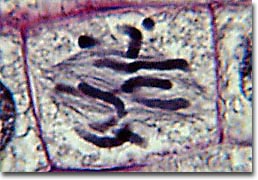
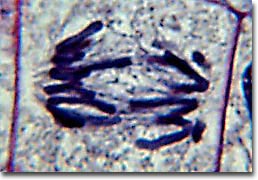
The first phase of mitosis is termed prophase. During prophase, the nuclear chromatin starts to become organized, condensing into the thick strands that eventually become chromosomes. The cytoskeleton (cytoplasmic microtubules) begins to disassemble and the mitotic spindle begins to form outside the nucleus at opposite poles of the cell.
Late prophase, or prometaphase commences with the disruption of the nuclear envelope, which is broken down into small membranous vesicles that closely resemble the endoplasmic reticulum, and which tend to remain visible around the mitotic spindle. During late prophase, the nucleolus disappears, and the chromosomes continue to condense, shortening and thickening. The microtubules of the mitotic spindle enter the nuclear region, while specialized protein complexes called kinetochore begin to form on each centromere. The kinetochores complexes become attached to spindle microtubules, which are now termed 'kinetochore microtubules'. Polar microtubules are not attached to centromeres and help to form and maintain the spindle structure along with astral microtubules, which remain outside the spindle.
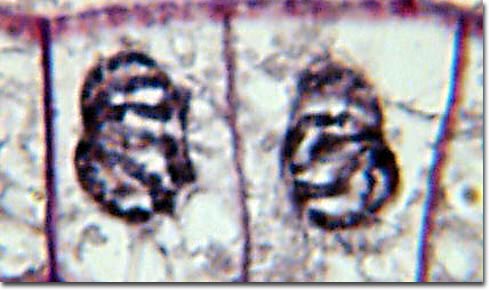
During metaphase, the chromosomes that are attached to the kinetochore microtubules begin to align along the metaphase plate, midway between the spindle poles. The kinetochore microtubules exert tension on the chromosomes prior to their separation during anaphase, which commences almost immediately after the metaphase chromosomes align at the metaphase plate. The two halves of each chromosome are pulled apart by the spindle apparatus during anaphase and migrate to the opposite spindle poles. Kinetochore microtubules shorten as the chromosomes are pulled toward the poles, while the polar microtubules elongate.
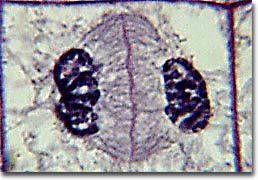
Anaphase is usually a rapid process that lasts only a few minutes. When the chromosomes have completely migrated to the spindle poles, the kinetochore microtubules begin to disappear, while the polar microtubules continue to elongate. This marks the junction between late anaphase and early telophase. Telophase which is the final stage in chromosome division during which the daughter chromosomes arrive at the spindle poles and are eventually redistributed into chromatin. The process of cytokinesis, where the cytoplasm is divided by cleavage, commences in late anaphase and continues through telophase.
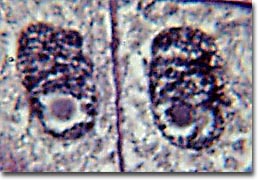
After complete separation of the chromosomes and their extrusion to the spindle poles, the nuclear membrane begins to reassemble around each group of chromosomes at opposite poles of the cell. Nucleoli also reassemble in the two newly forming cell nuclei.
The inherent dynamic instability of microtubules can be modified by the interactions with microtubule-associated proteins (MAPs) and microtubule-regulatory proteins. The best-characterized MAPs are MAP-1, MAP-2, and tau proteins. MAPs can bind to microtubules, increasing their stability. The activity of MAPs is tightly regulated by their phosphorylation state. Altered phosphorylation state of MAPs has been positively linked to the pathogenesis of Alzheimer’s disease.
Growth factor signals activate protein kinases that catalyze phosphorylation of tubulin-binding domains of MAPs, causing them to detach from microtubules. XMAP215 is a highly conserved 215 kDa MAP, which plays an important role in controlling microtubular dynamics during the cell cycle. XMAP215 stabilizes the plus ends of microtubules, promoting elongation and preventing catastrophic shrinkage. At the onset of mitosis, higher phosphorylation of XMAP215 increases microtubular instability, causing disassembly. During the final phase of mitosis, protein phosphatase activity predominates as the microtubule array characteristic of interphase is re-established.
Control of the Cell Cycle:
Eukaryotic cells alternate genome doubling (S-phase) with genome splitting (mitosis, M-phase) to generate daughter cells with an identical chromosomal complement. The cell cycle consists of a signal-controlled sequence of physiological states G1 → S → G2 → M, where non-mitotic phases are termed 'interphase'. Quiescent cells are said to be in phase G0, in which they are not participating in the cell cycle, but are metabolically active.
In a normal resting cell, intracellular signaling proteins and genes remain inactive unless activated by extracellular growth factors. When the normal resting cell is stimulated by an extracellular growth factor, signaling proteins and genes are activated and the cell proliferates. More than one hundred genes are specifically involved in cell cycle control, these are the so called CDC-genes (cell division cycle genes). One of these genes, designated CDC28 in Saccharomyces cerevisiae or CDK1 (cyclin dependent kinase 1) in humans, controls the first step in the progression through the G1-phase of the cell cycle, and is therefore also called "start". This gene encodes a protein member of the cyclin dependent kinase family (CDK). Half a dozen different CDK molecules have been found in humans. Cyclins are proteins formed and degraded during each cell cycle, so-named because the levels of cyclins vary periodically during the cell cycle. Cyclins bind to the CDK molecules, thereby regulating the CDK activity and selecting the proteins to be phosphorylated. Periodic protein degradation is an important general control mechanism of the cell cycle. Cyclins were conserved during evolution. Today around ten different cyclins have been found in humans. The levels of CDK-molecules are constant during the cell cycle, but their activities vary because of the regulatory function of the cyclins. CDK and cyclin together drive the cell from one cell cycle phase to the next. Levels of cyclins D (G1), E and A (S), and B and A (mitotic) fluctuate during the cell cycle, and binding of appropriate cyclins to the cyclin-dependent kinases (CDKs) stimulating phosphorylation and activation.
Controlled by cdc2 kinase, filaments of vimentin, desmin, and lamins disassemble prior to or early in mitosis then reassemble after cell division. Phosphorylation of serine residues in the N-terminal domain of lamin A and vimentin by cdc2 kinase induces the disassembly of intact filaments and prevents reassembly. Lamins a nuclear signal sequence and form a filamentous support inside the inner nuclear membrane. They are phosphorylated at the end of prophase and this causes them to disassemble simultaneous with dissolution of the nuclear envelope. After cell division, they are dephosphorylated just before the nuclei of the daughter cells form and lamin filaments reassemble around each set of chromosomes.
The cell cycle is a highly coordinated process that is controlled at multiple checkpoints along the pathway – the G1/S and G2/M transitions of interphase as well as anaphase of mitosis. These checkpoints are critical in preserving the fidelity of the genome. Alterations in the genes that control checkpoint processes are linked to a number of human malignancies including colon, breast, lung, kidney, brain and skin cancers.
In a cancer cell, mutation in a proto-oncogene that encodes an intracellular signaling protein (normally activated only by extracellular growth factors) creates an oncogene. The oncogene encodes an altered form of the signaling protein that behaves as though activated despite the absence of growth factor binding. That is, the malignant cell has escaped normal gene regulation and cell cycle control mechanisms and exhibits unchecked proliferation. Tables Regulatory Proteins Sequences Cell signaling Cell Adhesion Molecules Second Messengers Immune Cytokines Malignant Transformation Oncogenes Proto-oncogenes
Meiosis is controlled by similar factors to those that control mitosis.
In a healthy organism, cellular proliferation of tissues is normally balanced by cell death, which occurs by programmed apoptosis. Apoptosis is induced via the stimulation of several different cell surface receptors in association with activation of caspases (cysteinyl aspartate-specific proteases). Caspases can be activated by two main pathways: the death receptor pathway and the mitochondrial pathway.
Tables Apoptosis vs Necrosis Apoptosis Regulatory Proteins Sequences Cell signaling Phosphate-handling enzymes Malignant Transformation Oncogenes Proto-oncogenes
meiosis : mitosis : mitotic spindle : DNA replication • A • adhesion • C • cell membranes • cellular adhesion molecules • cellular signal transduction • centrioles • chemotaxis • chloroplast • cilia & flagella • communication • concentration gradients • cytokine receptors • cytoplasm • cytoskeleton • E • energy transducers • endoplasmic reticulum • endosomes • exosome • F • flagella & cilia • G • Golgi apparatus • GPCRs ♦ GTPases • H • hormones • I • ion channels • L • lysosome • M • meiosis • microtubules • mitosis • mitochondrion • molecular-switches • N • Nitric Oxide • neurotransmission • neuronal interconnections • nuclear membrane • nuclear pore • P • pinocytosis • proteasome • protein degradation • pumps • R • receptor proteins • receptor-mediated endocytosis • S • second messengers • signaling gradients • signal transduction • spindle • structure • T • transport • two-component systems • U • ubiquitin • V • vacuole • vesicle •
. Mitosis: An Interactive Animation . Mitosis interactive Java tutorial .

One cell division from single diploid parent cell produces two diploid daughter cells:
1. prophase
2. metaphase
3. anaphase
4. telophase
(diagram Stages of Mitosis : images : micrograph - mitotic cells : labelled micrograph : micrograph prophase metaphase anaphase : micrograph anaphase : micrograph telophase : micrograph, early prophase, late prophase, metaphase, early anaphase, late anaphase, telophase, daughter cells, resting : Movie illustration of mitosis : Mito Movie)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Mitosis allows cell nuclei to split, providing each daughter cell with a complete set of chromosomes during cellular division, while cytokinesis is division of the cytoplasm. Before replicating cells leave interphase, they pass through the synthesis or S phase in which each chromosome is duplicated and condensed to form two sister chromatids joined at the centromere. Centromeres contain a specific DNA sequence, and are crucial to segregation of the daughter chromatids during mitosis.
Mitogens, or somatomedins, are molecules that stimulates a cell to divide. Most mitogens are proteins, and they stimulate signal transduction pathways that utilize mitogen activated protein kinases. Mitogens include cytokines, growth factors, hormones, neurotransmitters, cellular stress proteins, and cell adhesion ligands.
The first phase of mitosis is termed prophase. During prophase, the nuclear chromatin starts to become organized, condensing into the thick strands that eventually become chromosomes. The cytoskeleton (cytoplasmic microtubules) begins to disassemble and the mitotic spindle begins to form outside the nucleus at opposite poles of the cell.
Late prophase, or prometaphase commences with the disruption of the nuclear envelope, which is broken down into small membranous vesicles that closely resemble the endoplasmic reticulum, and which tend to remain visible around the mitotic spindle. During late prophase, the nucleolus disappears, and the chromosomes continue to condense, shortening and thickening. The microtubules of the mitotic spindle enter the nuclear region, while specialized protein complexes called kinetochore begin to form on each centromere. The kinetochores complexes become attached to spindle microtubules, which are now termed 'kinetochore microtubules'. Polar microtubules are not attached to centromeres and help to form and maintain the spindle structure along with astral microtubules, which remain outside the spindle.
During metaphase, the chromosomes that are attached to the kinetochore microtubules begin to align along the metaphase plate, midway between the spindle poles. The kinetochore microtubules exert tension on the chromosomes prior to their separation during anaphase, which commences almost immediately after the metaphase chromosomes align at the metaphase plate. The two halves of each chromosome are pulled apart by the spindle apparatus during anaphase and migrate to the opposite spindle poles. Kinetochore microtubules shorten as the chromosomes are pulled toward the poles, while the polar microtubules elongate.
Anaphase is usually a rapid process that lasts only a few minutes. When the chromosomes have completely migrated to the spindle poles, the kinetochore microtubules begin to disappear, while the polar microtubules continue to elongate. This marks the junction between late anaphase and early telophase. Telophase which is the final stage in chromosome division during which the daughter chromosomes arrive at the spindle poles and are eventually redistributed into chromatin. The process of cytokinesis, where the cytoplasm is divided by cleavage, commences in late anaphase and continues through telophase.
After complete separation of the chromosomes and their extrusion to the spindle poles, the nuclear membrane begins to reassemble around each group of chromosomes at opposite poles of the cell. Nucleoli also reassemble in the two newly forming cell nuclei.
The inherent dynamic instability of microtubules can be modified by the interactions with microtubule-associated proteins (MAPs) and microtubule-regulatory proteins. The best-characterized MAPs are MAP-1, MAP-2, and tau proteins. MAPs can bind to microtubules, increasing their stability. The activity of MAPs is tightly regulated by their phosphorylation state. Altered phosphorylation state of MAPs has been positively linked to the pathogenesis of Alzheimer’s disease.
Growth factor signals activate protein kinases that catalyze phosphorylation of tubulin-binding domains of MAPs, causing them to detach from microtubules. XMAP215 is a highly conserved 215 kDa MAP, which plays an important role in controlling microtubular dynamics during the cell cycle. XMAP215 stabilizes the plus ends of microtubules, promoting elongation and preventing catastrophic shrinkage. At the onset of mitosis, higher phosphorylation of XMAP215 increases microtubular instability, causing disassembly. During the final phase of mitosis, protein phosphatase activity predominates as the microtubule array characteristic of interphase is re-established.
Control of the Cell Cycle:
Eukaryotic cells alternate genome doubling (S-phase) with genome splitting (mitosis, M-phase) to generate daughter cells with an identical chromosomal complement. The cell cycle consists of a signal-controlled sequence of physiological states G1 → S → G2 → M, where non-mitotic phases are termed 'interphase'. Quiescent cells are said to be in phase G0, in which they are not participating in the cell cycle, but are metabolically active.
In a normal resting cell, intracellular signaling proteins and genes remain inactive unless activated by extracellular growth factors. When the normal resting cell is stimulated by an extracellular growth factor, signaling proteins and genes are activated and the cell proliferates. More than one hundred genes are specifically involved in cell cycle control, these are the so called CDC-genes (cell division cycle genes). One of these genes, designated CDC28 in Saccharomyces cerevisiae or CDK1 (cyclin dependent kinase 1) in humans, controls the first step in the progression through the G1-phase of the cell cycle, and is therefore also called "start". This gene encodes a protein member of the cyclin dependent kinase family (CDK). Half a dozen different CDK molecules have been found in humans. Cyclins are proteins formed and degraded during each cell cycle, so-named because the levels of cyclins vary periodically during the cell cycle. Cyclins bind to the CDK molecules, thereby regulating the CDK activity and selecting the proteins to be phosphorylated. Periodic protein degradation is an important general control mechanism of the cell cycle. Cyclins were conserved during evolution. Today around ten different cyclins have been found in humans. The levels of CDK-molecules are constant during the cell cycle, but their activities vary because of the regulatory function of the cyclins. CDK and cyclin together drive the cell from one cell cycle phase to the next. Levels of cyclins D (G1), E and A (S), and B and A (mitotic) fluctuate during the cell cycle, and binding of appropriate cyclins to the cyclin-dependent kinases (CDKs) stimulating phosphorylation and activation.
Controlled by cdc2 kinase, filaments of vimentin, desmin, and lamins disassemble prior to or early in mitosis then reassemble after cell division. Phosphorylation of serine residues in the N-terminal domain of lamin A and vimentin by cdc2 kinase induces the disassembly of intact filaments and prevents reassembly. Lamins a nuclear signal sequence and form a filamentous support inside the inner nuclear membrane. They are phosphorylated at the end of prophase and this causes them to disassemble simultaneous with dissolution of the nuclear envelope. After cell division, they are dephosphorylated just before the nuclei of the daughter cells form and lamin filaments reassemble around each set of chromosomes.
The cell cycle is a highly coordinated process that is controlled at multiple checkpoints along the pathway – the G1/S and G2/M transitions of interphase as well as anaphase of mitosis. These checkpoints are critical in preserving the fidelity of the genome. Alterations in the genes that control checkpoint processes are linked to a number of human malignancies including colon, breast, lung, kidney, brain and skin cancers.
In a cancer cell, mutation in a proto-oncogene that encodes an intracellular signaling protein (normally activated only by extracellular growth factors) creates an oncogene. The oncogene encodes an altered form of the signaling protein that behaves as though activated despite the absence of growth factor binding. That is, the malignant cell has escaped normal gene regulation and cell cycle control mechanisms and exhibits unchecked proliferation. Tables Regulatory Proteins Sequences Cell signaling Cell Adhesion Molecules Second Messengers Immune Cytokines Malignant Transformation Oncogenes Proto-oncogenes
Meiosis is controlled by similar factors to those that control mitosis.
In a healthy organism, cellular proliferation of tissues is normally balanced by cell death, which occurs by programmed apoptosis. Apoptosis is induced via the stimulation of several different cell surface receptors in association with activation of caspases (cysteinyl aspartate-specific proteases). Caspases can be activated by two main pathways: the death receptor pathway and the mitochondrial pathway.
Tables Apoptosis vs Necrosis Apoptosis Regulatory Proteins Sequences Cell signaling Phosphate-handling enzymes Malignant Transformation Oncogenes Proto-oncogenes
meiosis : mitosis : mitotic spindle : DNA replication • A • adhesion • C • cell membranes • cellular adhesion molecules • cellular signal transduction • centrioles • chemotaxis • chloroplast • cilia & flagella • communication • concentration gradients • cytokine receptors • cytoplasm • cytoskeleton • E • energy transducers • endoplasmic reticulum • endosomes • exosome • F • flagella & cilia • G • Golgi apparatus • GPCRs ♦ GTPases • H • hormones • I • ion channels • L • lysosome • M • meiosis • microtubules • mitosis • mitochondrion • molecular-switches • N • Nitric Oxide • neurotransmission • neuronal interconnections • nuclear membrane • nuclear pore • P • pinocytosis • proteasome • protein degradation • pumps • R • receptor proteins • receptor-mediated endocytosis • S • second messengers • signaling gradients • signal transduction • spindle • structure • T • transport • two-component systems • U • ubiquitin • V • vacuole • vesicle •
. Mitosis: An Interactive Animation . Mitosis interactive Java tutorial .